What are we doing?

Shewanella & Pseudomonas
Our group is mainly using species of the genus Shewanella and Pseudomonas as model organisms to study various aspects of the bacteria in general. Both species are versatile motile gammaproteobacteria that can thrive in a wide range of environmental conditions. They have potential to be used in different bioremediation processes and applications, but have also been identified as commensal pathogens. The numerous sequenced species that were isolated from various environments make them well-suited models for studies on general microbial processes. All genetic tools are available for members of this genus.
Cell organization
Spatiotemporal organization is crucial for a number of fundamental cell processes such as cell division or motility in bacteria. We are particularly interested in processes located to the flagellar cell poles, e.g., synthesis of polar flagella and polar regulators, and the mechanisms which establish the polar identity.

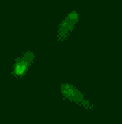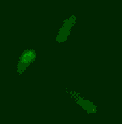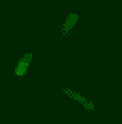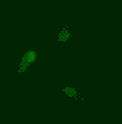
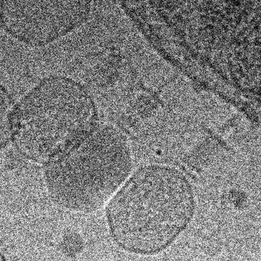
Phage biology
The viruses of bacteria, the phages, are extremely abundant and are thought to outnumber their hosts by a factor of ten. Over billions of years, phages and bacteria are engaged in a race-of-arms and affect each other and their populations in various ways we are only beginning to understand. The advent of advanced techniques in e.g. molecular biology, protein biochemistry, microscopy and next-generation sequencing and the identification of a huge number of novel phages has lead to a new wave of research on phages. We are interested in the molecular mechanisms underlying various aspects of phage-host interaction.

supported by the